| Authors | W. Gams & D. Hawksw. 1976 |
|---|---|
| Strain | 17035 |
| Classification | Hypocreales, Sarocladiaceae, Sarocladium |
| Culture collection | BCRC FU31124 |
| Detection frequency | Low |
| Accession number | LC461460 |
| Figure | 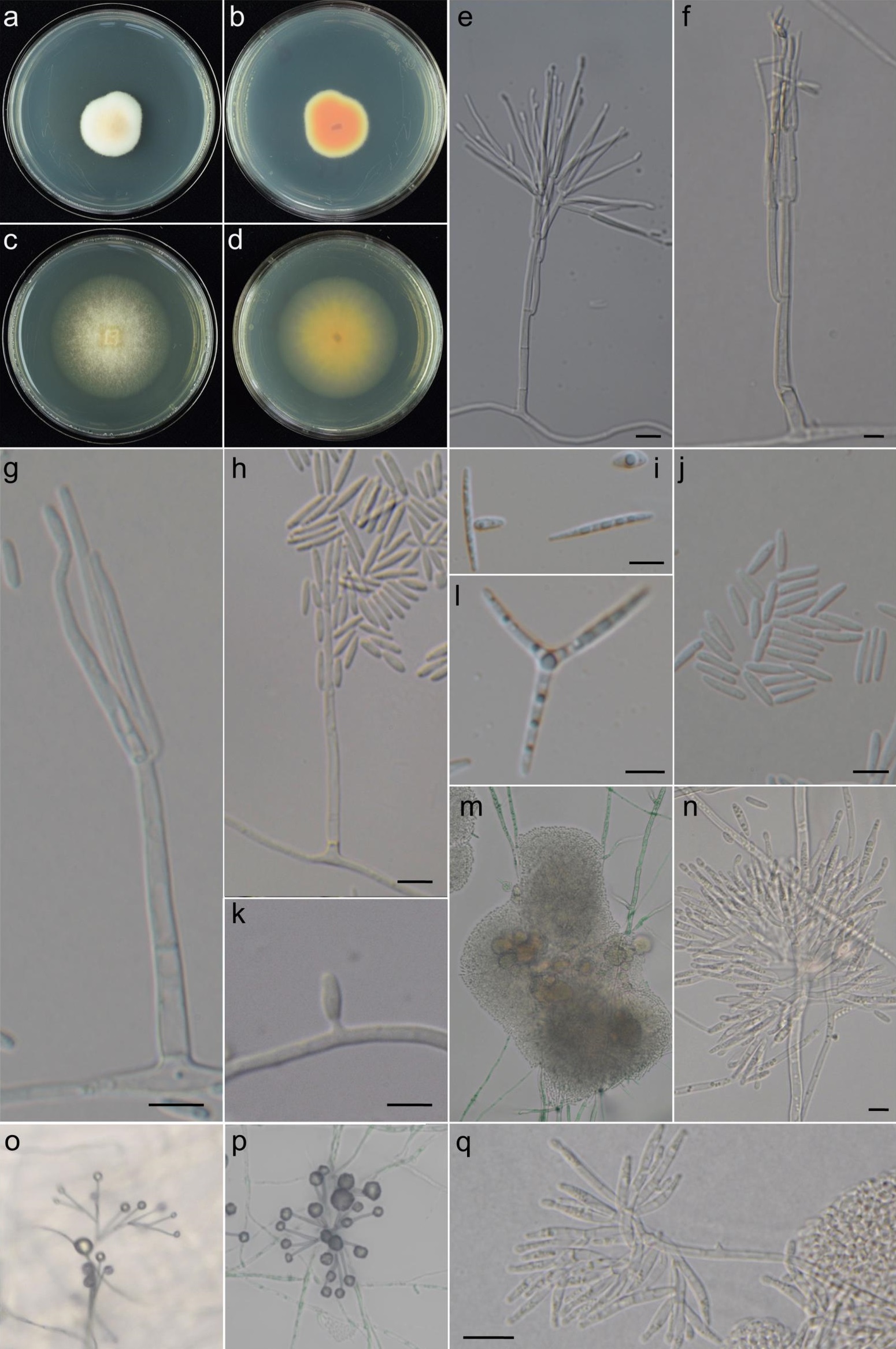 Fig. 1 Sarocladium attenuatum. a–d. Colony on PDA (a), with reverse in b and MEA (c), with reverse in d. e–h. Conidiophores and phialides, with conidia in g and h. i, l. Transformed and triradiate (three-armed) conidia. j. Conidia. k. Adelophialide with a conidium. m–n, q. Sporodochia., o–p. Phialides bearing liquid droplets consisting of conidial masses. (a–d from BCRC FU31118; e–f, j from strain SA 1- 5; g, m–n, q from strain SA 2-2; h, o–p from strain SA 3-3; i, l from strain SA 3-24; k from strain SA 3-36) (Bars = 5 μm) Fig. 1 Sarocladium attenuatum. a–d. Colony on PDA (a), with reverse in b and MEA (c), with reverse in d. e–h. Conidiophores and phialides, with conidia in g and h. i, l. Transformed and triradiate (three-armed) conidia. j. Conidia. k. Adelophialide with a conidium. m–n, q. Sporodochia., o–p. Phialides bearing liquid droplets consisting of conidial masses. (a–d from BCRC FU31118; e–f, j from strain SA 1- 5; g, m–n, q from strain SA 2-2; h, o–p from strain SA 3-3; i, l from strain SA 3-24; k from strain SA 3-36) (Bars = 5 μm) |
| Colonies | Colonies on PDA attaining 10-, 14-, 13-, and 7-mm diam. after 7 days at 24 °C, 28 °C, 32 °C, and 37 °C, respectively, elevated, wooly, white, sometimes becoming dark olive green when grown below 24 °C, margin entire, reverse salmon with white margin. Colonies on MEA attaining 16-, 17-, 18-, and 4- mm diam. after 7 days at 24 °C, 28 °C, 32 °C, and 37 °C, respectively, sparsely cottony, white-yellow at center, translu- cent at margin, reverse yellowish. |
| Sporodochia | Sporodochia present in some strains, consisting of short and densely aggregated branches bearing whorls of 9–18- μm-long and 1.5–2.5- μm-wide phialides. |
| Conidiophores | Conidiophores mononematous, macro- or micronematous, arising from sub- merged hyphae or aerial mycelium. Conidiophores reduced and bearing single phialides, simple and up to 70 μm high, terminating in 1–4 verticillately arranged phialides, repeatedly verticillately branched, where the central primary axis extends in growth with successive 1–3 lateral secondary axis arising along the primary axis or is determinate in growth and termi- nates in usually 2 and up to 4 verticillate branches, 1 or 2 of which being independent secondary new axis and the others of which being phialides, appearing monochasial, dichasial, or pleiochasial, the secondary axis developed in a similar manner as primary axis, with or without tertiary axis. Conidiophore axes usually strongly adpressed, turning to slightly divergent toward the distal part, occasionally with additional branches arising vertically from the lateral sides of secondary or tertiary axes. |
| Conidiogenous cells | Phialides cylindrical, 33–45 × 1–2 μm, terminally forming solitary or in whorls of 2–4, or intermixed with conidiophore axes to form a whorl of 2–4 at the branching points, amounting to maximum of 40 individuals on each conidiophore. Short lateral adelophialides sometimes present. |
| Conidia | Conidia hyaline, one-celled, non-guttulate, cylindrical with rounded ends, 3.5–8 × 1.0–2.0 μm (length/width ratio 3.1–8.5), forming a slimy mass on top of phialides. Additional conidia, if present, usually guttulate, ovoid to cylindrical with sharply pointed ends, var- iable in length, up to 25 μm long, sometimes triradiate (three-armed). |
| Note | There has been a debate about the status of S. attenuatum. Gams and Hawksworth (1975) established this species, and distinguished it from S. oryzae in having verticillate branching pattern of conidiophores and longer sized conidia. Bridge et al. (1989) observed the continuum of conidial size between S. oryzae and S. attenuatum, and suggested that S. attenuatum is a synonym of S. oryzae. Bills et al. (2004) sequenced the ITS and LSU regions of ex-type material of S. oryzae (CBS 180.74) and S. attenuatum (CBS 399.73) and reported that these two strains had 100% identity in ITS and LSU regions, and consequently supported the synonymous treatment of Bridge et al. (1989). However, Summerbell et al. (2011) re-sequenced the ITS region of CBS 399.73 and especially stated that they obtained a sequence (not deposited in database) different from the sequence of Bill et al. (2004) (AY566995) in 6 base pairs and two gaps. Summerbell et al. (2011) then concluded that the status of S. attenuatum required further studies. Giraldo et al. (2015), by comparing ITS, LSU. and ACT sequences, indicated that the similarity between S. oryzae isolates CBS 180.74 and S. attenuatum CBS 399.73 was in the range between 98.4% and 98.8%. They acquired an ITS sequence (HG965027) of CBS 399.73, differing from Bill et al. (2004) (AY566995), conforming to the analysis of Summerbell et al. (2011). Apparently there was a mistake made by Bill et al. (2014). The ITS sequence, AY566995, should not be from S. attenuatum, but from S. oryzae instead. However, Giraldo et al. (2015) preferred to maintain S. attenuatum as a synonym of S. oryzae.
In addition to the cylindrical shape in conidia, elongated cylindrical, ovoid and tri-radiate shapes were occasionally observed in S. attenuatum. Elongated conidia have also been reported by Bridge et al. (1989) and Brady (1980), whereas ovoid and tri-radiate conidia were here observed for the first time. We found that these additional kinds of conidia were formed only occasionally and always dislodged and encountered on the colony surface. These conidia were always associated with free water. The intergradation in conidial sizes found with S. attenuatum and S. oryzae by Bridge et al. (1989) was likely an artifact of including transformed conidia in the measurements. Accordingly, when the size range of conidia is based on those freshly produced in masses on phialides, a distinction could be clearly made in conidial size between S. attenuatum and S. oryzae. Sporodochia, composed of aggregated conidiophores, were observed for the first time. Phialides produced from sporodochia are clearly shorter, and differ between S. attenuatum and S. oryzae. They are subulate in S. attenuatum and flask-shaped in S. oryzae. Bill et al. (2004) noted that S. attenuatum CBS 399.73 and S. oryzae CBS 180.74 can produce green colonies. During the growth tests, we found that temperatures below 24°C were crucial to induce the green pigment. However, only a few strains of S. attenuatum were able to produce green pigment in this condition. Pathogenicity was confirmed by artificial inoculation with 3 strains (BCRC FU31124, BCRC FU31127, BCRC FU31118). Grain discoloration, unfilled grains, and lesions on leaf sheaths were observed on inoculated rice plants. |
| Pathogenicity | This species causes rice sheath rot disease. |
| Specimens examined | Taiwan, Miaoli County, rice grains, Sep 2018, Jie-Hao Ou, 18076 |
| ITS | CTCCAACCCATTGTGAACATACCTATCGTTCCCTCGGCGGGACCAGCGCGCGGCTGCCTCAGGGCTCCGGGCGTCCGCCGGGGACAGCCAAACTCGAATTTTGCAGTGATTCTCTGAGGGGCGAGAGCCCGAAAACAAAATAAATCAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCGGCACTCCGGCGGGCATGCCTGTCCGAGCGTCATTTCAACCCTCAGGAGCACCCTTCGGGGCGCACCTGGCGCTGGGGATCAGCGGCCCTCGCGGCCCCTGTCCCCCAAATTCAGTGGCGGTCGCGCCGCAGCCTCCCCTGCGTAGTAGCACAACCTCGCACCGGAGAGCGGAACGACCACGCCGTGAAACCCCCAACTTTCCAAGGTTGACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAGC |
| LSU | ACCCGCTGAACTTAAGCATATCAATAAGCGGAGGAAAAGAAACCAACAGGGATTGCCTCAGTAACGGCGAGTGAAGCGGCAACAGCTCAAATTTGAAATCTGGCCGCAAGGTCCGAGTTGTAATTTGCAGAGGATGCTTTTGGCGAGGTGCCTTCCGAGTTCCCTGGAACGGGACGCCATAGAGGGTGAGAGCCCCGTACGGTTGGACCACCGAGCCTCTGTAAAGCTCCTTCGACGAGTCGAGTAGTTTGGGAATGCTGCTCTAAATGGGAGGTGTACGTCTTCTAAAGCTAAATACCGGCCAGAGACCGATAGCGCACAAGTAGAGTGATCGAAAGATGAAAAG |
| ACT | GTCCTGCTCACCGAGGCGCCCATCAACCCCAAGTCCAACCGTGAGAAGATGACCCAGATTGTCTTCGAGACCTTCAACGCCCCCGCATTCTACGTCTCTATCCAGGCCGTTCTGTCTCTGTACGCCTCTGGTCGTACCACCGGTATCGTTTTGGACTCTGGTGATGGTGTCACTCACGTTGTCCCCATCTACGAGGGTTTCGCCCTCCCCCACGCCATTGCTCGTGTCGACATGGCTGGCCGTGATCTTACTGATTACCTCATGAAGATCCTTGCTGAGCGTGGTTACACCTTCTCCACCACCGCCGAGCGTGAAATCGTTCGTGACATCAAGGAGAAGCTCTGCTACGTCGCCCTCGACTTCGAGCAGGAGATCCAGACCGCCGCCCAGTCCTCCAGCCTCGAGAAGTCTTACGAGCTTCCTGACGGCCAGGTCATCACCATCGGCAACGAGCGATTCCGCGCCCCTGAGGCTCTCTTCCAGCCCTCCGTCCTCGGTCTCGAGAGCGGTGGTATCCACGTCACCACTTTCAACTCCATCATGAAGTGCGATGTCGACGTCCGAAAGGATCTCTACGGCAACATTGTCATGGTATGTTTTAGGTCAAACCAGAGATCTGTGACGATTGAAAACTAACAATGACCTAGTCTGGTGGTACCACTATGTACCCTGGTCTCTCCGACCGTATGCAGAAGGAGATCACTGCTCTTGCTCCTTCTTCCATGAAGGTCAAGATCATTGCTCCTCCGGAGCGCAAGTACTCTGTC |
| TUB | AGGACGTTGTCAACGAGCTCAGCCCCCTCAGTGTAGTGACCTTTGGCCCAGTTGTTGCCAGCACCGGACTGGCCGAAAACGAAGTTGTCAGGGCGGAAAAGCTGACCGAAGGGACCAGCACGGACGGCATCCATGGTACCGGGCTCGAGATCGACGAGGACAGCGCGAGGAACATACTTGTTGCCGGAGGCCTGAAGAGGCGGTCAGTGGGTCCATAGCAAGCAAGAGACGCGTTGTGAGCATCTCTCAGTCATATCCAACCACGAACCTCGTTGAAGTAGACGCTCATGCGCTCGAGCTGGAGGTCGGAGCTGCCATTGTAGATACCATTGCTGTCGAGACCGTGCTCGCCGGAGATGGTCTGCCAGAAAGCAGCACCAACTTGGTTGCCCTAGATCGAAAACATGATGAGCCCCGCACATGAAGTCGAGATGGGAGCTATTGCAGCTACTTACACACTGGCCGGTCTGGAGGTGAATCTATTTGCAAAAGAAAAAGACATGGTTAGCACGTCATTTCTCTCTCAAAGCTTCATCAAATGTGTTGACACGCCACATGGCGATATCCCAAAGTCTCGGTGGTACCCCGCCACGAGTTTAGGGGTAACAGCGGTTTTGCTGGCAACAAAGGAACGCGTCGCGCGGGGGTTTCGAGGCTCGTGGCGCATGGGGGGGAGGAGGAATGGAACTCACAATC |